![]()
How did viruses evolve? Are they a streamlined form of something that existed long ago, or an ultimate culmination of smaller genetic elements joined together?
The evolutionary history of viruses represents a fascinating, albeit murky, topic for virologists and cell biologists. Because of the great diversity among viruses, biologists have struggled with how to classify these entities and how to relate them to the conventional tree of life. They may represent genetic elements that gained the ability to move between cells. They may represent previously free-living organisms that became parasites. They may be the precursors of life as we know it.
We know that viruses are quite diverse. Unlike all other biological entities, some viruses, like poliovirus, have RNA genomes and some, like herpesvirus, have DNA genomes. Further, some viruses (like influenza virus) have single-stranded genomes, while others (like smallpox) have double-stranded genomes. Their structures and replication strategies are equally diverse. Viruses, do, however, share a few features: First, they generally are quite small, with a diameter of less than 200 nanometers (nm). Second, they can replicate only within a host cell. Third, no known virus contains ribosomes , a necessary component of a cell's protein-making translational machinery.
To consider this question, we need to have a good understanding of what we mean by "life." Although specific definitions may vary, biologists generally agree that all living organisms exhibit several key properties: They can grow, reproduce, maintain an internal homeostasis , respond to stimuli, and carry out various metabolic processes. In addition, populations of living organisms evolve over time.
Do viruses conform to these criteria? Yes and no. We probably all realize that viruses reproduce in some way. We can become infected with a small number of virus particles — by inhaling particles expelled when another person coughs, for instance — and then become sick several days later as the viruses replicate within our bodies. Likewise we probably all realize that viruses evolve over time. We need to get a flu vaccine every year primarily because the influenza virus changes, or evolves, from one year to the next (Nelson & Holmes 2007).
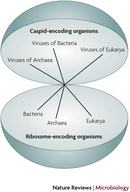
Viruses do not, however, carry out metabolic processes. Most notably, viruses differ from living organisms in that they cannot generate ATP . Viruses also do not possess the necessary machinery for translation , as mentioned above. They do not possess ribosomes and cannot independently form proteins from molecules of messenger RNA . Because of these limitations, viruses can replicate only within a living host cell. Therefore, viruses are obligate intracellular parasites. According to a stringent definition of life, they are nonliving. Not everyone, though, necessarily agrees with this conclusion. Perhaps viruses represent a different type of organism on the tree of life — the capsid-encoding organisms, or CEOs (Figure 1; Raoult & Forterre 2008).
There is much debate among virologists about this question. Three main hypotheses have been articulated: 1. The progressive, or escape, hypothesis states that viruses arose from genetic elements that gained the ability to move between cells; 2. the regressive, or reduction, hypothesis asserts that viruses are remnants of cellular organisms; and 3. the virus-first hypothesis states that viruses predate or coevolved with their current cellular hosts.

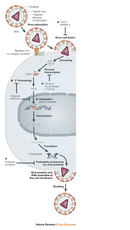
According to this hypothesis, viruses originated through a progressive process. Mobile genetic elements, pieces of genetic material capable of moving within a genome , gained the ability to exit one cell and enter another. To conceptualize this transformation , let's examine the replication of retroviruses, the family of viruses to which HIV belongs.
Retroviruses have a single-stranded RNA genome. When the virus enters a host cell, a viral enzyme , reverse transcriptase , converts that single-stranded RNA into double-stranded DNA. This viral DNA then migrates to the nucleus of the host cell. Another viral enzyme, integrase , inserts the newly formed viral DNA into the host cell's genome. Viral genes can then be transcribed and translated. The host cell's RNA polymerase can produce new copies of the virus's single-stranded RNA genome. Progeny viruses assemble and exit the cell to begin the process again (Figure 2).
This process very closely mirrors the movement of an important, though somewhat unusual, component of most eukaryotic genomes: retrotransposons. These mobile genetic elements make up an astonishing 42% of the human genome (Lander et al. 2001) and can move within the genome via an RNA intermediate. Like retroviruses, certain classes of retrotransposons, the viral-like retrotransposons, encode a reverse transcriptase and, often, an integrase. With these enzymes, these elements can be transcribed into RNA, reverse-transcribed into DNA, and then integrated into a new location within the genome (Figure 3). We can speculate that the acquisition of a few structural proteins could allow the element to exit a cell and enter a new cell, thereby becoming an infectious agent. Indeed, the genetic structures of retroviruses and viral-like retrotransposons show remarkable similarities.
In contrast to the progressive process just described, viruses may have originated via a regressive, or reductive, process. Microbiologists generally agree that certain bacteria that are obligate intracellular parasites, like Chlamydia and Rickettsia species , evolved from free-living ancestors. Indeed, genomic studies indicate that the mitochondria of eukaryotic cells and Rickettsia prowazekii may share a common, free-living ancestor (Andersson et al. 1998). It follows, then, that existing viruses may have evolved from more complex, possibly free-living organisms that lost genetic information over time, as they adopted a parasitic approach to replication.
Viruses of one particular group, the nucleocytoplasmic large DNA viruses (NCLDVs), best illustrate this hypothesis. These viruses, which include smallpox virus and the recently discovered giant of all viruses, Mimivirus, are much bigger than most viruses (La Scola et al. 2003). A typical brick-shaped poxvirus, for instance, may be 200 nm wide and 300 nm long. About twice that size, Mimivirus exhibits a total diameter of roughly 750 nm (Xiao et al. 2005). Conversely, spherically shaped influenza virus particles may be only 80 nm in diameter, and poliovirus particles have a diameter of only 30 nm, roughly 10,000 times smaller than a grain of salt. The NCLDVs also possess large genomes. Again, poxvirus genomes often approach 200,000 base pairs, and Mimivirus has a genome of 1.2 million base pairs; while poliovirus has a genome of only 7,500 nucleotides total. In addition to their large size, the NCLDVs exhibit greater complexity than other viruses have and depend less on their host for replication than do other viruses. Poxvirus particles, for instance, include a large number of viral enzymes and related factors that allow the virus to produce functional messenger RNA within the host cell cytoplasm .
Because of the size and complexity of NCLDVs, some virologists have hypothesized that these viruses may be descendants of more complex ancestors. According to proponents of this hypothesis, autonomous organisms initially developed a symbiotic relationship. Over time, the relationship turned parasitic, as one organism became more and more dependent on the other. As the once free-living parasite became more dependent on the host, it lost previously essential genes. Eventually it was unable to replicate independently, becoming an obligate intracellular parasite, a virus. Analysis of the giant Mimivirus may support this hypothesis. This virus contains a relatively large repertoire of putative genes associated with translation — genes that may be remnants of a previously complete translation system. Interestingly, Mimivirus does not differ appreciably from parasitic bacteria, such as Rickettsia prowazekii (Raoult et al. 2004).
 A black arrow points from the blue bacterial cone to the red Archaeal cone. A question mark above the arrow suggests there is uncertainty regarding the evolutionary relationship between the modern bacterial and archaeal viruses." />
A black arrow points from the blue bacterial cone to the red Archaeal cone. A question mark above the arrow suggests there is uncertainty regarding the evolutionary relationship between the modern bacterial and archaeal viruses." />
The progressive and regressive hypotheses both assume that cells existed before viruses. What if viruses existed first? Recently, several investigators proposed that viruses may have been the first replicating entities. Koonin and Martin (2005) postulated that viruses existed in a precellular world as self-replicating units. Over time these units, they argue, became more organized and more complex. Eventually, enzymes for the synthesis of membranes and cell walls evolved, resulting in the formation of cells. Viruses, then, may have existed before bacteria, archaea , or eukaryotes (Figure 4; Prangishvili et al. 2006).
Most biologists now agree that the very first replicating molecules consisted of RNA, not DNA. We also know that some RNA molecules, ribozymes, exhibit enzymatic properties; they can catalyze chemical reactions. Perhaps, simple replicating RNA molecules, existing before the first cell formed, developed the ability to infect the first cells. Could today's single-stranded RNA viruses be descendants of these precellular RNA molecules?
Others have argued that precursors of today's NCLDVs led to the emergence of eukaryotic cells. Villarreal and DeFilippis (2000) and Bell (2001) described models explaining this proposal. Perhaps, both groups postulate, the current nucleus in eukaryotic cells arose from an endosymbiotic-like event in which a complex, enveloped DNA virus became a permanent resident of an emerging eukaryotic cell.